
SUBUNIT COMPOSITION OF ATP SYNTHASE
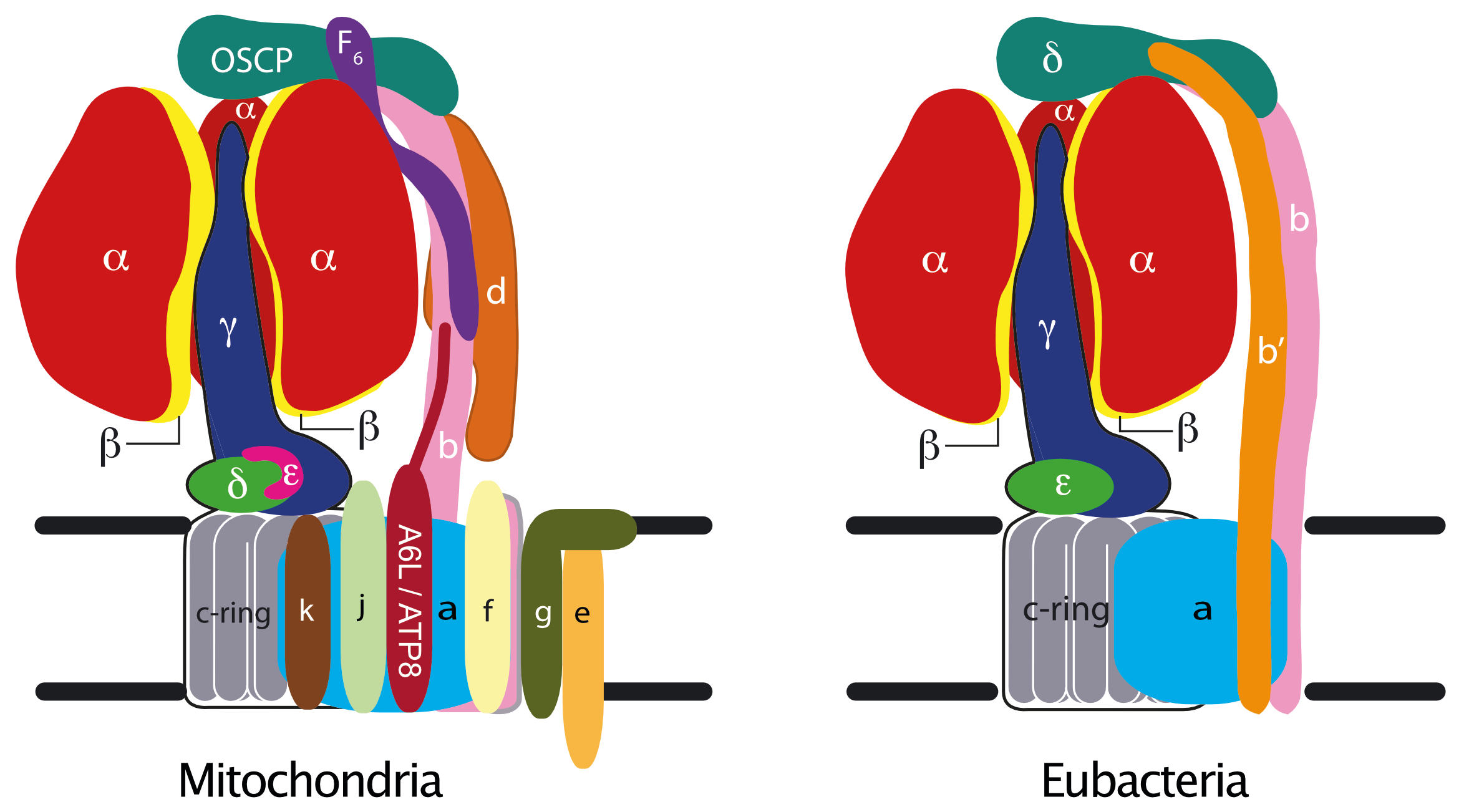
ATP synthase has two major structural parts known as F1 and Fo linked by the peripheral and central stalks. The F1 catalytic domain of the mitochondrial enzyme is a globular assembly of five different proteins, α, β, γ, δ and ε with the stoichiometry 3:3:1:1:1 [1]. The three α-subunits and the three β-subunits are arranged alternately around a central α-helical coiled-coil in the γ-subunit. The γ-subunit protrudes from α3β3-subcomplex and the δ- and ε-subunits are associated with its foot. The foot interacts with a ring of c subunits in the membrane domain. The Fo domain is an assembly of single copies of each of the ATP6 (or a), b, c, e, f, g, and ATP8 (or A6L) subunits, all of them embedded in the inner mitochondrial membrane [2] [3] [4]. Mammalian enzymes contain additional supernumerary subunits j and k (formerly known as 6.8 PL (6.8 kD proteolipid) and DAPIT (diabetes associated protein in insulin sensitive tissue)) [5] [6], and fungal enzymes, supernumerary subunits j, k and l [7]. Subunit l is closely related to subunit k. The peripheral stalk (made of single copies of subunits OSCP, b, d and F6), the a-subunit and the associated supernumerary subunits constitute the enzyme’s "stator", against which the rotor, made of the c-ring and subunits γ, δ and ε, turns. Passage of protons through a pathway between the c-ring and subunit a releases energy to drive the clockwise rotation of the rotor (as viewed from the membrane) during ATP synthesis. The c8-ring found in the bovine and human enzymes probably persists throughout metazoans [8] [9]. S. cerevisiae and Pichia angusta have c10-rings [10]. The functions of subunits e, f, g, DAPIT and 6.8PL are becoming less obscure as the resolution and accuracy of cryo-em structures of dimeric ATPases improve. [11].
The subunit compositions of bacterial and chloroplast ATP synthases are simpler than those from mitochondria [12] [13] [14], and the complexes are monomeric. The F1 catalytic domain consists of the α3β3-subcomplex plus the central stalk made of single copies of subunits γ and ε. The bacterial or chloroplast ε-subunit is the orthologue of the mitochondrial δ-subunit [15]. The bacterial δ-subunit is the orthologue of the mitochondrial OSCP [15] and forms part of the peripheral stalk which contains either two identical copies of the b-subunit (as in Escherichia coli for example), or a single copy of the b-subunit plus one copy of a related b’-subunit, each with a single transmembrane α-helix (as in chloroplasts and Paracoccus denitrificans, for example). Mitochondrial and bacterial peripheral stalk subunits have no obvious relationships between their sequences, and the peripheral stalks are the most divergent regions among the common domains of the ATP synthases. In mycobacteria, the δ- and b-subunits are fused. The number of c-subunits in the c-rings in bacterial ATP synthases varies from 9-17 [16] [17] [18].
REFERENCES
- Walker JE, Fearnley IM, Gay NJ, Gibson BW, Northrop FD, Powell SJ, Runswick MJ, Saraste M & Tybulewicz VL (1985)
Primary structure and subunit stoichiometry of F1-ATPase from bovine mitochondria.
J Mol Biol 184, 677-701 - Fearnley IM & Walker JE (1986)
Two overlapping genes in bovine mitochondrial DNA encode membrane components of ATP synthase.
EMBO J 5, 2003-2008 - Walker JE, Lutter R, Dupuis A & Runswick MJ (1991)
Identification of the subunits of F1F0-ATPase from bovine heart mitochondria.
Biochemistry 30, 5369-5378 - Collinson, IR, van Raaij, MJ, Runswick, MJ, Fearnley, IM, Skehel, MJ, Orriss, GL, Miroux, B & Walker, JE (1994)
ATP synthase from bovine heart mitochondria: in vitro assembly of a stalk complex in the presence of F1-ATPase and in its absence.
J Mol Biol 242, 408-421 - Chen R, Runswick MJ, Carroll J, Fearnley IM & Walker JE (2007)
Association of two proteolipids of unknown function with ATP synthase from bovine heart mitochondria.
FEBS Lett 581, 3145-3148 - Meyer B, Wittig I, Trifilieff E, Karas M & Schägger H (2007)
Identification of two proteins associated with mammalian ATP synthase.
Mol Cell Proteomics 6, 1690-1699 - Liu S, Charlesworth TJ, Bason JV, Montgomery MG, Harbour ME, Fearnley IM & Walker JE (2015)
The purification and characterization of ATP synthase complexes from the mitochondria of four fungal species.
Biochem J 468, 167-175 - Watt IN, Montgomery MG, Runswick MJ, Leslie AGW & Walker JE (2010)
Bioenergetic cost of making an adenosine triphosphate molecule in animal mitochondria.
Proc Natl Acad Sci U S A 107, 16823-16827 - Walpole TB, Palmer DN, Jiang H, Ding S, Fearnley IM & Walker JE (2015)
Conservation of complete trimethylation of lysine-43 in the rotor ring of c-subunits of metazoan adenosine triphosphate (ATP) synthases.
Mol Cell Proteomics 14, 828-840 - Stock D, Leslie AG & Walker JE (1999)
Molecular architecture of the rotary motor in ATP synthase.
Science 286, 1700-1705 - Päivärinne H & Kainulainen H (2001)
DAPIT, a novel protein down-regulated in insulin-sensitive tissues in streptozotocin-induced diabetes.
Acta Diabetol 38, 83-86 - Foster DL & Fillingame RH (1979)
Energy-transducing H+-ATPase of Escherichia coli. Purification, reconstitution, and subunit composition.
J Biol Chem 254, 8230-8236 - Foster DL & Fillingame RH (1982)
Stoichiometry of subunits in the H+-ATPase complex of Escherichia coli.
J Biol Chem 257, 2009-2015 - Walker JE, Saraste M & Gay NJ (1984)
The unc operon. Nucleotide sequence, regulation and structure of ATP-synthase.
Biochim Biophys Acta 768, 164-200 - Walker JE, Runswick MJ & Saraste M (1982)
Subunit equivalence in Escherichia coli and bovine heart mitochondrial F1F0 ATPases.
FEBS Lett 146, 393-396 - Meier T, Polzer P, Diederichs K, Welte W & Dimroth P (2005)
Structure of the rotor ring of F-Type Na+-ATPase from Ilyobacter tartaricus.
Science 308, 659-662 - Matthies D, Preiss L, Klyszejko AL, Müller DJ, Cook GM, Vonck J & Meier T (2009)
The c13 ring from a thermoalkaliphilic ATP synthase reveals an extended diameter due to a special structural region.
J Mol Biol 388, 611-618 - Preiss L, Langer JD, Yildiz Ö, Eckhardt-Strelau L, Guillemont JEG, Koul A & Meier T (2015)
Structure of the mycobacterial ATP synthase Fo rotor ring in complex with the anti-TB drug bedaquiline.
Sci Adv 1, e1500106